

文 | 吴乐旻
加州大学伯克利经济学博士,曾任教于北大经济学院,新书《富种起源》即将出版
企业的边界,反映了社会对隔断度的优化。在竞择中脱颖而出的社会,是在“企业内部学习”和“阻断模仿以保持多样化”之间取得有效平衡的社会。
英国著名畅销书作家Matt Ridley 在 2010 年 TED 全球大会上曾经做过一个演讲,题目很性感,叫“When Ideas Have Sex”(当思想有性繁殖)↵。
有性繁殖和思想有什么关系呢?
我们知道,无性繁殖是由母体的一部分直接形成新个体。而有性繁殖允许参与繁殖的多个个体组合他们的基因。
比如,人类两性会各拿己方的一部分染色体切割重组成儿女的染色体。这种高度随机的切割重组是基因多样性除突变以外另一大重要源泉。
相比之下,无性繁殖缺少有性繁殖那种多个个体间交流基因的机会(只存在少数例外)。

基因的交流,配合上自然选择,可以把一个种群内较有适应性的基因组合起来——即使这些基因最早出现在许多不同的个体身上。而无性繁殖只能依靠孤独的演化主脉自己去累积所有所需的突变。
Matt Ridley 想说,社会能够快速进步,就是依靠人类思想这种类似于有性繁殖的特征。
如果思想失去了“性”——失去了人与人之间、社会和社会之间思想上的交流和重组——文明就容易丧失多样性,固步自封,陷入停滞。
这段演讲的视频在 TED官网上已有 250 多万的观看量。Ridley 的这一思想也并不是他的原创。类似的观点早已在各种媒体上流传,为人们所熟悉。可能有些读者一看到本文的题目,When Ideas Have Sex,就猜我也要宣扬这一思想。
但恰恰相反,我想说的是:有性繁殖和生物多样性间的关系并不是Ridley所理解的那样。其中的一波三折,处处情理之中,又无不是意料之外,很值得欣赏玩味。
边域种化与间断平衡说
演化生物学家Steven Stanley↵曾在1975 年的一篇论文↵中指出,有性繁殖相对于无性繁殖,其实在很大程度上是限制了生物的多样性,而不是扩充了多样性。
Stanley的思想,源自20 世纪最伟大的演化生物学家之一 Ernst Mayr (1954)↵的边域种化论(Allopatric Speciation Theory)↵。

Mayr 认为,生物多样性的来源和达尔文的判断不一样。新物种不是来自母种群的种系渐变,而是来自与母种群隔离开的小规模子种群的急剧变化。这一学说后来被Eldredge & Gould (1977)↵总结为间断平衡说。
种系渐变论和间断平衡说各自假想了迥然不同的物种演化分叉图(图 1)。种系渐变论下,物种分裂的枝杈是 y 字形的(我特地用小写 y)。各物种在其存续期内始终保持缓慢的演化。依靠这种渐变,种群隔离后两个分开的种群就会向着不同的方向各自演化出去。
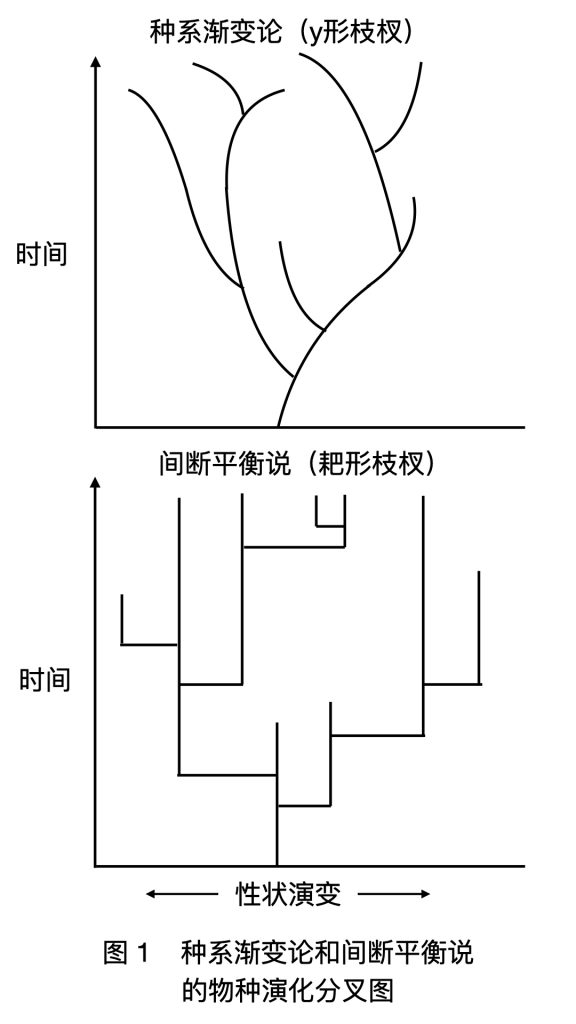
而间断平衡说的枝杈是耙形的。子种群一旦与母种群隔离,就摆脱了母种群的基因流,打破了多基因间互动所造成的自平衡稳态(Homeostasis)。于是,子种群的基因迅速变化,直到变化积累到足够程度,实现与母种群间的生殖隔离,就形成了新的物种,亦即发生了“种化”(speciation)。
但种群在稳定之后,物种形态将保持高度的稳定。在间断平衡说看来,基因本有巨大的变化潜力,却受制于基因流的自平衡稳态,在母种群中发挥不出来;只有在边域种化打破平衡的短暂瞬间才凶光毕露。新的物种几乎完全来自边域种群的隔离、漂移和对特殊生态位的适应。
基因流的自平衡稳态
Stanley 的边域种化理论有一个关键概念:基因流的自平衡稳态。它是什么意思呢?
众所周知,基因的表达一般要牵扯到许多不同的基因。在一个稳定的、有着丰富基因库的母种群里,各基因在该种群所栖息的环境里接近最优的适应度,靠的是各种基因间的配合。
一条基因能在这个母种群里占据优势,靠的是许多辅助基因时不时地窜到一个个个体的身上,与这条基因配合,产生功效。母种群通过内部的基因交流共享一个基因流(gene-flow);任何一条基因都得放到整个基因流中才能体现它的价值。
这一特征所带来的结果是,有性繁殖的大规模母种群一般演化极为缓慢。每当有新基因出现的时候,初来乍到的新基因因为缺乏与现有基因流的配合,很难单枪匹马胜过“人脉宽广”的等位旧基因。
但是,当母种群分离出一个很小的子种群到一个孤立的环境(比如小岛)时,这个子群的基因多样性就萎缩了。
旧的等位基因将失去很大一部分本来可以支持自己的辅助基因——它的“人脉”断了,落在母种群里了。这时候,新基因就比较容易胜过旧的等位基因。
基因流的隔绝、基因库的残缺,意味着基因之间的组合、竞争要重新洗牌。这种情况下,哪怕这个小岛的环境和母种群所在的环境差不多,物种的形态也容易发生大的变化。

隔离小种群的存活率远不如大种群。但万一它们活了下去,并在新的生态位扩张自己,将来一旦和母种群的地理隔绝终结,这个子种群有可能会杀回老家,甚至代替曾经的母种群。
这样的变化表现在化石记录上,就是古生物学家们曾经感到困惑的间断平衡现象——一个新的物种,几乎没见到任何过渡形态,怎么就突然出现了?甚至,一个上百万年形态稳定的物种怎么突然就变成另一个物种了?
其实,过渡形态是存在的,但过渡形态都发生在那个短暂的隔离初始阶段里那个狭小的隔离环境里,即使有化石留存,化石被古生物学家发现的概率也微乎其微。这些问题往往被神创论者用来攻击进化论,但当你明白间断平衡说之后,你就会觉得:要是化石证据不是这样的,那才奇怪呢。
在Mayr (1954) 的基础上,Stanley(1975a)提出了一个叫做“物种竞择”的理论框架。
他说自然选择有两层。
一层是我们所熟悉的发生在种群内个体间的竞择。在这个层面上,有性繁殖的重组能带来基因多样性,有益于物种的存衍。
另一层竞择发生在物种之间。物种可以通过子种群隔离“繁殖”出新的物种,又可以通过隔离子种群的种化实现物种的“突变”。隔离越频繁,新物种就越多,就越有利于物种所在门类整体的存衍和繁荣。
Eldredge & Gould(1977)后来用物种竞择的思想探讨过小哺乳动物的相对优势:因为小哺乳动物的个体活动范围较小,更容易发生子种群被隔绝开的事件,因此在物种多样性上胜过大哺乳动物。

Stanley (1975b) 则用物种竞择的思想探讨有性繁殖的相对优势。
Stanley 设问:大家都说有性繁殖能通过基因重组来获得多样性,还说这种增加的多样性能帮助有性繁殖物种避开偶发的环境危机,实现更长久的物种存续;但是,如果比较有性繁殖和无性繁殖物种的存续时间,却会发现有性繁殖并不具备优势。这是为什么?
另外,无性繁殖一般出现在简单生物,比如原核生物中,而有性繁殖一般出现在复杂生物中。这又是为什么?
有性繁殖的宏观优势
Stanley 感到,答案就在Mayr 的思想里。站在个体角度,有性繁殖能够分享基因流里突变自其它个体的优秀基因。但站在整个物种的角度上说,有性繁殖的基因重组却是一支相反的力量:它通过 Mayr 所说的基因流自平衡稳态,把基因变化的潜能锁了起来,使物种反而无法适应多样化的环境。
所以光看单个物种,有性繁殖并不比无性繁殖拥有更大的多样性优势,当然也就谈不上物种存续时间的优势。
不过,我这么说并不严谨。有性繁殖下的物种可以用生殖隔离来定义。但无性繁殖下每个个体只是略有变化的克隆体而已,没有生殖,谈何生殖隔离,没有生殖隔离,谈何物种,所以无性繁殖下的物种定义是模糊而主观的。把有性繁殖下生殖隔离意义上的一个物种,拿来和无性繁殖下分类学家靠看图说话来界定的物种作比较,缺乏可比性。好在 Stanley 的这个意思,读者们应该是都能明白的。

Stanley 的这个逻辑,如果拿文化传播做比,就好比说:许多个体间相互借鉴学习,站在个体立场上固然是丰富了知识的多样性。但也因为这种借鉴学习,造成个体间的同化,以致丧失了群体的多样性。
对物种的存续来说,性状的多样性可能是最最重要的一个维度。如果有性繁殖的基因流稳态反而损害了基因多样性,“性”在演化意义上不就是一种自杀吗?那为什么世界上还有性这一回事呢?有性繁殖能够在复杂生物中占据主导,一定有其它的机制来弥补多样性。
你可能已经想到:有性繁殖实现多样性,与其说是靠着Matt Ridley 所讴歌的基因重组,不如说是靠着Mayr 所发现的边域种化。基因重组对个体来说是一支有利于适应环境的力量,但对物种来说却是一支锁死变化、造成停滞的力量。真正造成多样性的,是隔离,隔离,隔离。和 Ridley 所说的完全相反!
边域种化所带来的当然不是母种群、母物种的延续。它所服务的是物种之上更高等级生物门类的整体存续。
隔离越频繁,新物种就越多,就越有利于物种所在门类整体的存衍和繁荣。
有性繁殖从隔离产生种化。这一层级的物种竞择反而跟无性繁殖的竞择类似:新物种是旧物种的一个变异克隆(可以把有性繁殖生物的“物种”类比为无性繁殖生物的“个体”)。只不过,这种克隆方式的变异程度要比无性繁殖从母代到子代间的变化大得多。
就单一物种来说,无性物种对环境适应范围的扩张速度比有性物种更快,但它快不过有性繁殖物种所在的大门类通过种化所实现的整体性的环境适应范围的扩张速度。
如果用生物对环境的适应范围的扩张速度来度量生物的环境适应能力,那么粗糙地说,无性物种的适应能力比有性物种强,但比不过有性物种所在门类的环境适应能力。
为什么无性繁殖物种适应范围的扩张比不上有性繁殖门类的种化速度呢?
因为有性繁殖门类下新物种的形成靠的是隔离小种群的边域种化。隔离种群摆脱了基因流的封锁,就能尽情施展有性繁殖的重组优势,把不同个体身上的好基因汇集起来,快速种化,以适应所陷入的新环境。而无性繁殖只能靠单脉谱系去积累优势基因,适应的速度就不如有性繁殖的边域种化。所以,不能说 Ridley 完全没有道理,但他的道理,他的喻体,只在漫长的物种存续史中电光火石的一小段里才算成立。
基因多样性的好处是能让物种在灭绝事件中存活。灭绝事件就像横在某段生态位上的一堵墙(图 2)。有性繁殖如果没有种化,基因互动重组所导致的基因流自锁只会让有性繁殖更难适应环境变化。所以图 2中有性繁殖的树状图里每个枝杈都是笔挺挺的——它的扩张速度比左侧无性繁殖图里的枝杈更小。但是凭借种化,有性繁殖在整体上反而拥有了比无性繁殖更优秀的“绕墙能力”。从个体优势,到物种劣势,又到门类优势,有性繁殖和多样性的关系可谓峰回路转,一波三折。
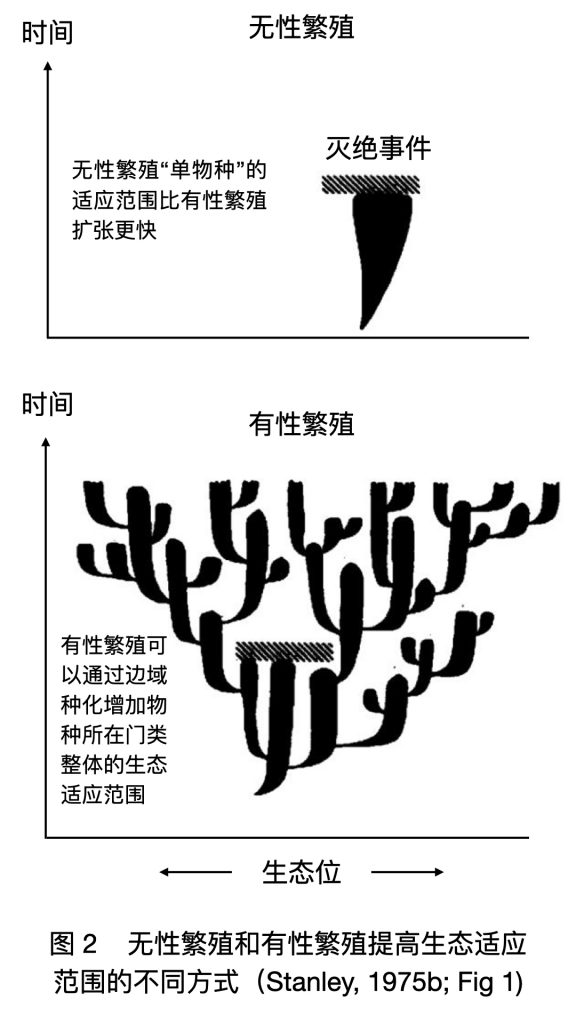
那为什么复杂生物比简单生物更倾向于有性繁殖呢?Stanley 的解释是,当一类生物本身就能适应很多不同的环境,灭绝率本来就很低时,它不用有性繁殖也能好好地存续下去。在图 2中,这就好比一个物种的枝杈(横向适应)本来就已经很宽了,一般的墙拦不住它,它光靠无性繁殖那几下子就能活得好好的。但那些适应环境比较狭窄,很容易因环境变化而灭绝的生物,搞无性繁殖就太容易灭绝了,能存活下来的门类就得是那些掌握性手段的生物。
简单生物的适应面本来就广,所以原核生物还保留着无性繁殖。复杂生物本来适应面窄,所以复杂生物多用有性繁殖,或者说在有了性之后,生物演化打开了从简单向复杂蔓延的大门。
Stanley 的假说当然不是有性繁殖优势的唯一解释。关于性的起源,还有许许多多解释。我对这个解释感兴趣。一是它和我们通常认可的有性繁殖增加物种适应范围这一观点截然相反,很有趣。二是它对人类社会内制度、技术、文化、思想的传播规律也有很多呼应。下面我就谈谈这个理论对社会科学的启发。
文明演化中的边域种化
人类文明也有文化交流带来的铁索连环现象,文化的均一化会妨碍族群对本地环境的充分适应。
假使有一个中心地带的农耕文明形成了一个文化区。受中心地带影响,文化区中的人民即使在边缘更适合放牧的地区也选择了种地;至于边缘之外那些没法种地、只适合放牧的地区,甚至都没有人去占领。
突然之间,一个边缘地区与中心地带隔绝了——比如地理上产生了阻隔或政治上发生了分裂——失去了文化交流(迷因流分流了)。边缘地区的农民一开始肯定很不适应。但慢慢地,文化开始改变,开始适应当地的环境。
人民渐渐从耕作转向了本地环境更适宜的放牧。转向之后,这些边区人民赫然发现,外面更广大的无人区也可以放牧啊,于是占领了那块地方。这个原本人口稀疏、很不起眼的边缘群落,就成了一大片地方的主人,人口在新生活方式下的繁盛远胜于往昔。
他们自立为一个新的民族。甚至靠征服,反过来完成了对农耕民族旧文化的代替。于是,历史书(文明的化石)上就表现为长期的文化稳定后发生了一场间断平衡式的突变。
征服农耕民族后,如果农耕区更大,这个征服民族又会接受文化上的同化。他们被同化之后,反而不再适应放牧生活,又会造成放牧区人口密度的下降。接着,也许是新一轮的隔绝、外拓、征服、蜕变。
农耕-游牧的故事可能有点俗套。我们还可以想想英国工业革命的事情。英国孤悬海外,这个地理位置特别适合发生文化迷因的突变(迷因可以理解为思想的基因)。海峡造成阻隔,允许英国文化更自由地去适应本地环境。当航海时代到来的时候,英国就能捷足先登。

英国王室的土地曾经一度横跨英吉利海峡。英国王室的心思甚至还更偏欧洲大陆一点。如果这样一个领土安排继续下去,英国在航海时代还能像后来那样成功吗?
英国在航海上的成功,如果类比到Mayr 的物种演化上,不就是隔绝之后,实现本土化适应,并以此开拓、占领一个外围的新生态位吗?占领这个生态位以后,英国人殖民到美洲,发展起工业文明,最后工业文明作为一种文化技术迷因还席卷了全世界,包括中国。以至于未来的考古学家眼中,中国在几十年间也经历了一场间断平衡,文明的样貌在缓慢变化了上千年后,突然之间就变了个模样。
企业的迷因流
企业管理中也有铁索连环现象。
企业就像一个种群,种群内部有着频繁的基因交流。大企业能容纳更多的人才、更多的想法,这些想法在企业内部有性繁殖,为整个企业所分享。这是大企业的优势。
但与此同时,企业文化的均一化也桎梏了企业对环境的充分适应。我们经常说船小好调头,这背后的道理就有点像母种群基因流的自稳定平衡,是一种 organizational homeostasis:大企业牵一发而动全身,不好轻易改革,连原理都和 Mayr (1954) 一致。
针对组织均一化造成的适应度缺陷,通常的应对方式是在大企业里划分出灵活的、有一定自主权的事业部和子公司来,主动割裂大企业内部的文化基因流。这种割裂能够切断母体基因流,允许子体更充分地适应特殊小环境,占领本来不得不放弃的边缘生态位。
人类企业要比物种灵活。一大原因是,人类企业的内部是高度分工的。一个1 万人的大企业,具体到一个工种,一个工序,可能只有 30 个人在做,和小种群(小企业)人数差不多。这 30 个人所进行的创新,尤其是技术创新,往往是独立于大企业其它特征的,是能够适应这道工序的“特殊环境”的。在这种按照工序定义的生态位上,大企业的适应度不见得有多弱。
大企业所不能适应的生态位,一般是市场生态位,而不是工种生态位。而之所以大企业难以适应市场生态位,是因为开拓市场所涉及的企业文化基因有着更复杂繁密的 Epistasis(基因互动)。所以,在涉及到市场开拓的业务上,大企业仍有划分子公司的必要。

但是子公司的独立性是有限的。它们能帮大企业在一些边缘生态位敲敲边鼓,却很难承担开天辟地、“形成新物种”的使命。也就是说,对一个经济体来说,大企业的体内隔离终究不能彻底代替小企业的演化价值。和有性繁殖的边域隔离物种一样,小企业的预期寿命远逊于大企业,但小企业船小好调头,繁荣的小企业群体是社会多样性的宝库。
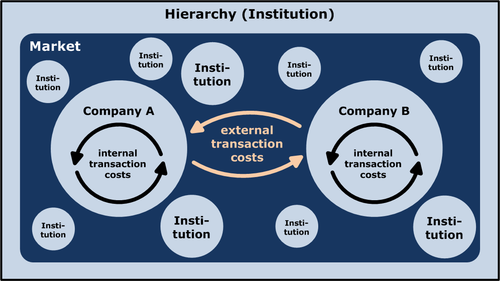
从企业基因交流的角度,我们甚至可以对科斯的市场-企业边界之问,补充一个答案。企业的边界,反映了社会对隔断度的优化。在竞择中脱颖而出的社会,是在“企业内部学习”和“阻断模仿以保持多样化”之间取得有效平衡的社会。这不是企业和企业在竞争,而是社会和社会在竞争,是由制度竞争筛选下来的“最优隔断度”。
以迷因滞性来定义制度
让我们再把视线投向国家。人类对模仿别国的技术是很起劲的,但是对模仿别国的制度是很消极的。是因为制度本身很难选择很难改吗?那倒不是。就和边域种化一样,一群移民扎根新大陆时,他们对制度选择往往是非常开放的,会涌现出大量精彩的设计(比如北美的清教徒移民社会)。

显然,制度基因存在高度的 Mayr 式的“基因互动”(异位显性),牵一发而动全身。所以,一个大的“母种群”搞制度改革很难。大种群要改制,往往需要全局式的 Big Push。而小种群利用基因锁链的短暂破缺,就能迅速变化(不过,人类凭借主观能动性,以及权力结构的金字塔特点,有时的确能实现big push,比如彼得大帝的改革、日本的明治维新)。

人类对制度改革的保守,不仅是因为克服异位显性的艰难,也在很大程度上出于各文化天然的保守心理——改革滞性。制度受制于竞择,大家都知道。但不太容易注意到的是,对制度改革的保守心理同样也受制于竞择。如果人类族群像学习技术一样热衷于搬用异族宪纲,对族群上层的文明区整体而言,岂不是丧失了制度基因的多样性?这样的文明区,最终也容易走向衰亡。
再说一层更玄的:人类对制度的定义也受制于竞择。人类文化有那么多潜在的基因:技术、文化、制度。这些都是看不见摸不着的思想,ideas。它们间的概念区别是哪里来的?
有没有可能,在所有的思想中,有些类型的思想从族群存衍的角度是适宜学习的,内部交流越多越好;而有些类型的思想涉及到大规模的基因互动,容易限制族群适应环境的能力,或者造成基因流自稳定平衡,锁死整体的多样性,所以不适宜“拿来主义”?

有没有可能,人类自然而然地把前一类型的思想称作技术,而后一种思想,按照它不适宜拿来的程度,称作文化和制度。于是,先有人类天性中的体用之别,再有制度与技术之分——“彼之技术为用,我之制度为体”,学一个,不学一个;再从这技术、制度的区分中,人类自然而然地衍生出组织的概念,对组织属性——制度——采取保守态度?
这个思想实验天马行空,比起制度和技术的通常定义来说似乎舍近求远,但我的目的不在于挑战通俗的概念,而在于探寻一种新的思考角度。这个思考角度不是完全没有道理的。
想象一种纯粹的技术,按理说应该是积极学习模仿的对象,但现在社会演化上出了个小bug。某个民族偏偏对这项技术避之不及。他们会怎么找理由呢?是不是把它归结到神神鬼鬼的文化制度上?龙脉啊,风水啊,摄魂啊……到头来,一种思想,到底是技术还是文化制度,本来不就是灵活的吗?技术和制度的区别,从社会演化的角度,可能源于看似毫不相关的学习滞性。而这个学习滞性受长期演化影响,体现了社会的“最优隔断度”。
所以,到底是保守定义了组织,还是组织定义了保守?
从应然的角度说,制度和文化的适应范围窄。为了防止在灭绝事件中全军覆没,平时就不能老汇流,汇到后来,多样性没了。站在更高层看去,还不如适当减少汇流,增加分流,最大化制度的多样性。
而技术型基因不同。相对于技术型障碍,技术的适应范围本来就宽,汇流之后宽度还能增加。所以放开汇流,增加的多样性一般是超过损失的多样性的。结果,人类社会的竞择可能呈现出一种该稳定的稳定(制度和文化)、该动荡的动荡(技术)这样一种格局。就好比生物演化,有的走上了有性繁殖,有的走上了无性繁殖。

当然,在生物基因和思想迷因之间做类比,要懂得节制,把握类比的界限。迷因和基因的不同点至少包括下面两项:
第一,迷因不是随机的。选择什么迷因组合起来,往往有人的主观能动性在里头。
第二,迷因即使“种化”了,跨种的杂交还是很普遍。因此,文化迷因不仅在个体竞择上像有性繁殖,而且和有性繁殖不同,它的上层“物种竞择”不像无性繁殖,而仍然像有性繁殖。这个意义上,思想迷因比大自然里的有性物种更富有性特征。所以,我在上面的讨论里,没有把迷因的“隔离”处理成非此即彼的 0-1 变量,而将之视为有程度之分的一个连续变量。
后记:这篇随笔改写自 2021 年 3 月份我的两篇学术自修日记。我是一个社科工作者,因研究需要,阅读了几篇演化生物学有关间断平衡说的经典文献,当时写了几篇日记,当做日常功课而已。
时隔半年,蒙DC不弃,要在他的新杂志上发表此文。但隔行如隔山;这几篇生物学文献在后续几十年的发展,我也未曾跟踪,一时深恐误人子弟,贻笑大方。
犹豫之际,读到日记末尾引用王羲之的话: “仰观宇宙之大,俯察品类之盛,所以游目骋怀,足以极视听之娱,信可乐也”。想到此文面世则能与万千读者分享这种快乐,终于还是不揣冒昧,甘效野人献曝的厚颜。如果我的理解有错,还请读者海涵,指正。
主要参考文献
Gould, S.J. and Eldredge, N., 1972. Punctuated equilibria: an alternative to phyletic gradualism. Models in paleobiology, pp.82-115.
Gould, S.J. and Eldredge, N., 1977. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology, pp.115-151.
Mayr, Ernst (1954). “Change of genetic environment and evolution” In J. Huxley, A. C. Hardy and E. B. Ford. Evolution as a Process. London: Allen and Unwin, pp. 157-180.
Stanley, S. M. 1975a. A theory of evolution above the species level. Proc. Nath. Acad. Sci. 72:646-650.
Stanley, S. M. 1975b. Clades versus clones in evolution: why we have sex. Science. 190:382-383.
责任编辑:DC